
alinooova

Подскажите пожалуйста логику рассуждений в данном задании и как из графиков понять концентрацию АТФ? чем больше протов в апопласте, тем больше их будет проходить через АТФазу?
Спасибо за объяснение
Подскажите пожалуйста логику рассуждений в данном задании и как из графиков понять концентрацию АТФ? чем больше протов в апопласте, тем больше их будет проходить через АТФазу?
Спасибо за объяснение

“Кислый рост” индуцируется ауксином при конкретном ее концентрации. Модель выглядит примерно так: Ауксин связывается с H+ протонным насосом, и протоны закачиваются в клеточную стенку, и пониженный pH “активирует” белки режущие связи между полисахаридами в клеточной стенке, и соответственно давление клеточный стенки снижается, и клетка удлиняется. Видимо сахароза является источником АТФ, который необходим для закачки протонов водорода. Уже этими выводами можем убрать варианты А, С и D. Так как роль ауксина является активация протонного насоса (хотя не уверен на счет того, что функция ауксина в этой модели только эта), то и вариант E отпадает. Вариант B самый подходящий. Думаю суть вопроса смог объяснить. А на счет концентрации атф, нам ее ведь не нужно определять? Информация об обработке сахарозой уже достаточна. И совместная “работа” ауксина и сахарозы создает оптимальные условия для удлинения, что и можем наблюдать по графику.
P.s Индол-уксусная кислота - это ауксин
upd: сахароза для котранспорта протонов водорода, проситите(


I. кДНК это типо ДНК копии мРНК. Она имеет такую же последовательность что и мРНК, только рнк нуклеотиды замененые на днк нуклеотиды. Если говорить о праймерах, то они определают направленность молекулы.

II. Так как на электрофорезе мы видим несколько разных длин кДНК, то можно предположить что они были получены с помощью альтернативного сплайсинга.
III. Самый длинный фрагмент на геле приблизительно равен 500bp, а это значение можно получить только если все пять экзонов захваченных двумя праймерами были бы в мРНК.
IV. Если бы делали ПЦР используя начальную ДНК, то все интроны были бы в составе. Выходит 3598bp.

Привет Арунна,
Утверждение 1 неверно так как мы видим рост с отрицательного конца на графике (после точки D)
Утверждение 2 верно, так как тредмиллинг обозначает рост с одного конца и сокращение с другого конца. Между В и D плюс конец выше нуля, а минус конец ниже, что означает что один конце растет, а другой сокращается – определение тредмиллинга.
Утверждение 3 верно, так как минус конец пересекает ось икс гораздо позже чем плюс конец. Критическая концентрация полимеризации это концентрация при которой филамент начинает расти (то есть пересекает ось икс) Плюс конец начинает расти около 0.2 микромоль актина, а минус конец около 0.8 микромоль.
Утверждение 4 неверно, так как скорость роста плюс конца выше чем скорость сокращения минус конца при концентрация 0.5 микромоль. Это означает, что размер меняется.
Утверждение 5 неверно, так как эти белки работают на промежуточных филаментах, а не актиновых.

Вот схемы для двух видов аутосплайсинга:

Интересно то, что если в этом процессе участвует сплайсосома, реакция похожа на 2 тип:


А как здесь играет роль структура РНК? Есть ли закономерность того, что лассо движется медленнее ? То есть, в задаче РНК по структуре лассо движется медленнее. Почему так?
Похожую картину можно наблюдать при работе с плазмидными векторами. Если одновременно запустить в гель кольцевую молекулу ДНК и разрезанную по одному сайту, то можно увидеть примерно такую картину:
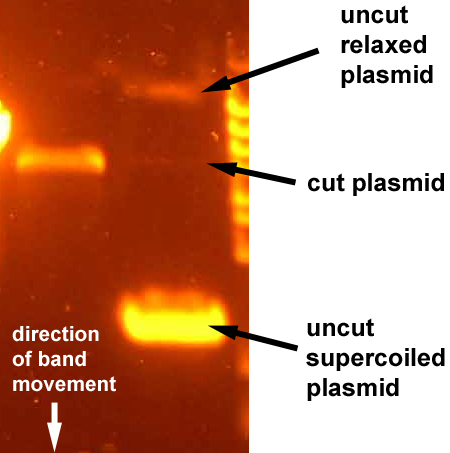
Тут даже чуть больше разнообразия: релаксированная плазмида (в ней скорее всего есть однонитевой разрыв), разрезанная плазмида, суперскрученная плазмида. Их положение на геле зависит от того, как они сопротивляются движению по агарозному гелю.

Можно оценить их компактность, но предсказывать лучше ничего не стоит. В этом задании говорится далеко не про стандартную форму РНК, так что я делал все выводы только на сравнении и имеющихся данных.


Электрохимическая сила — это комбинация электрического потенциала мембраны (то как заряды распределны по двум сторонам мембраны) и градиента концентрации. Я лично знаю вот такую формулу которая суммирует эти два параметра:
Первая часть показывает влияние градиента концентрации, вторая — влияние электрического потенциала мембраны. Для калия градиент больше, однако из-за того, что он, в основном, находится в месте, где отрицательный заряд (внутренняя сторона мембраны), то этот заряд его удерживает и общая электрохимическая сила будет не такой значительной.
Для натрия хоть градиент концентрации и меньше, заряд внешней стороны положительный и он, наборот, толкает натрий внутрь клетки. Получается, что у натрия происходит суммирование эффекта градиента концентрации и мембранного потенциала.
1 ответ
Для третьего.
В уравнении каждая концентрация иона умножена на проницаемость мембраны для него (p_{Na} для натрия). Чем больше это значение, тем большее влияние на общее значение будет оказывать концентрация этого иона. К примеру если для натрия это будет 100, а для калия будет 0.1, то даже самый огромный градиент концентрации калия не будет значительно влиять на потенциал - можешь подставаить в уравнение и проверить.
то есть, у калий эти два параметра(градиент концентрация и электрический потенциал мембраны) противоположны, и их суммация приводит к незначительной электрохимической силе? А вот, например, в потенциал покое обычного нейрона все так же, но внутри клетки наоборот больше отрицательного заряда, при этом внутри больше калий. В этом случае так же два параметра у калий противоположны. Здесь тоже её электрохимическая сила меньше будет?

По закону Харди-Вайнберга p^2+2pq+q^2=1
где q – частота рецессивного аллеля (вероятность встретить рецессивный ген), p – частота доминантного аллеля.
Среди мужчин могут быть два варианта: здоровый X^HY (частота p) или болеющий X^hY (частота q).
Среди женщин могут быть три варианта: здоровая гомозиготная X^HX^H (частота p^2), здоровая гетерозиготная A.K.A носительница рецессивного гена X^HX^h (частота 2pq) и болеющая гомозиготная X^hX^h(частота q^2).
Тогда частота болеющих мужчин и женщин = 0.5q + 0.5q^2 = 0.12 = 12 \%
где 0.5 обозначает равную долю мужчин и женщин в популяции.
Решив уравнение q^2+q-0.24=0 получим q = 0.2 (второе значение q – отрицательное, что не является физически возможным).
Подставив q=0.2 в p^2+2pq+q^2=1 получим p = 0.8 (опять же, второе значение p не является физически возможным).
Если взять из популяции здоровых мужчину (p) и женщину (p^2+2pq), то рождение болеющего мальчик зависит только от гетерозиготной женщины (от отца Х-сцепленный рецессивный ген не сможет получить), которую можно встретить с вероятностью в 2pq.
В добавок к этому нужно учесть что вероятность рождения болеющего мальчика у конкретной пары X^HY и X^HX^h равна 1/4.
Тогда вероятность рождения болеющего мальчика у здоровой пары будет равна
\frac{1}{4} \cdot \frac{2pq}{p^2+2pq} = \frac{1}{4} \cdot \frac{2 \cdot 0.8 \cdot 0.2}{0.8^2+2 \cdot 0.8 \cdot 0.2} = \frac{1}{12}= 0.0833 \approx 8.3 \%
P.S. Для решения таких задач советую хорошенько обдумать, то как вероятность фенотипа зависит от вероятности нахождения X-сцепленного рецессивного гена для каждого пола.
1 ответ
Не надо ли нам здесь находить вероятность того, что отец не болен?
То есть, мы взяли частоту женщин только среди женщин. Если бы мы нашли бы частоту встречаемости гетеризигот женщин во всей популяции, в чем была бы наша ошибка?
2 ответа
В задаче уже говорится что родители здоровы, а если они здоровы то рецессивный ген может передаться мальчику только от матери.
Понял, рақмет
Объяснять график я начну с того, что предположу что ты знаешь что такое уравнение Михаэлиса-Ментен (но вот краткое напоминание):
Оно не очень удобно в плане построения графика и его оценки, поскольку график этого уравнения будет выглядеть как гипербола:
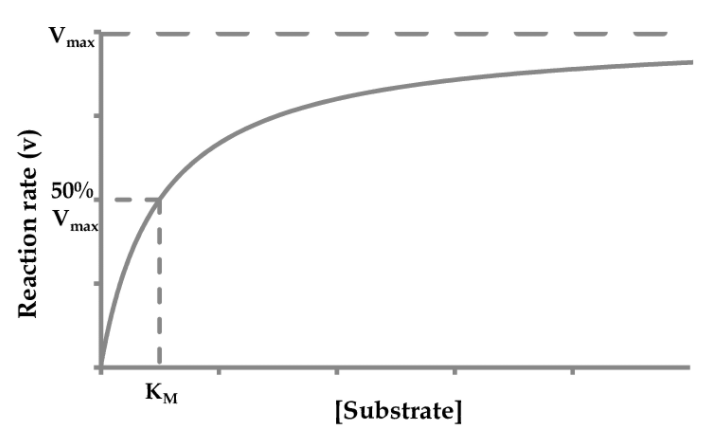
График стал таким:
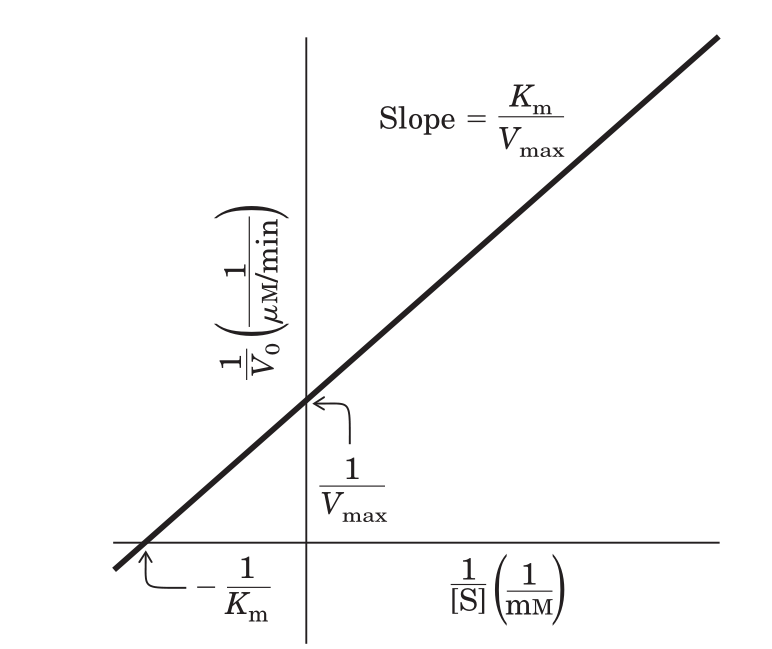
Смотрим на график в задании.
I. Тамифлю увеличивает Км - конкурентный ингибитор (уменьшается значение 1/Km)
II. Реленза - уменьшает Vmax (увеличивается 1/Vmax) - неконкурентный
III. Тамифлю конкурентный ингибитор - связывается с ферментом только когда активный сайт свободен
IV. Реленза неконкурентный ингибитор - связывается как с ферментом так и с ES комплеком.
V. В последнем дана скорость, но не дана концентрация субстрата и фермента, по графику без этого нельзя определить скорость.
Извините, я вот здесь не понял 1)как бесконкурентные ингибиторы заставляют Km снижаться ?
2) У нас в неконкурентном снижается Vmax, но не Km. Если допустим у нас 100 ферментов, и для 0.5Vmax нужно 50 субстратов, и Km = 50, но при неконкурентном инг. свободных снижается на 70, то значить ли это, что Km снизилось до 35, или она осталось 50? Или тут нужно смотреть на Km по другому ?
1)Как вообще выглядит реакция для бесконкурентных ингибиторов:

Теперь что такое Км:
По сути Км обратно зависим от сродства фермента к субстрату. Мне кажется на эту проблему стоит смотреть со стороны химического равновесия. Поскольку ингибитор связывается с ES, концентрация ES уменьшается и равновесие реакции смещается в сторону образования ES - прямая реакция от E + S => ES ускоряется. Соответственно больше ферментов связывают субстрат.
2) Да здесь Vmax уменьшилсь и соответственно 1/2 Vmax тоже стала меньше, при этом чтобы достигнуть половины от максимума с ингибитором и без него требуется одинаковое количество субстрата.
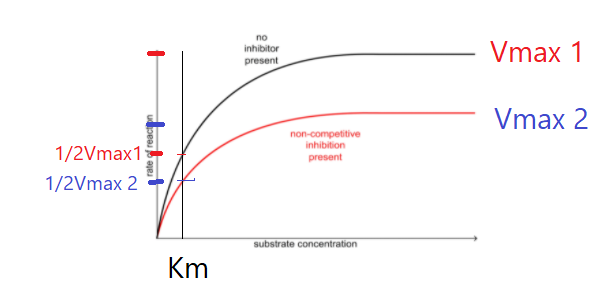
То есть, это примерно так рабоатет ? В неконкурентном ингибировании хоть и понижается максимальная средняя скорость, именно нужное количество субстрата для его достижения такой же остается. Это связано с тем, что нек.ингибитор не влияет на аффиность фермента. Но даже если он не влияет на афинность, часть оставшиеся связанным с ферментом ингибиторы не дает субстрату связаться с ферментом, хоть субстрат и растет. Поэтому она не дсотигает vmax. А вот в конкурентном ингибиторе , чем больше становится субстрата, ингибитор может разрушить связи ферментом и дать место субстрату, поэтому когда очень много субстрата ни один ингибитор не остается связанным с ферментом. Поэтому при конкурентном инг. скорость может достичь до Vmax. Но при этом так как нам нужно убрать ингибиторов(обратимые) нужно больше усилий, больше субстрат, больше Km ??
Да, вообще действие неконкурентных ингибиторов можно рассматривать как простое уменьшение концентрации фермента - ингибиторы связываются с аллостерическим сайтом и инактивируют фермент, с этого момента он вообще в остальных реакциях не участвует(пока обратно не распадется комплекс EI) соответственно концентрация рабочего Е падает.
В случае с конкурентными ингибиторами тоже примерно так как ты описал. Но правда ингибитор не просто может разрушить связь с ферментом, он это делает постоянно - там в реакции стрелка в две стороны, что значит что постоянно образуются ES, EI комплексы и постоянно они диссоциируют, просто все это находится в равновесии. И тут дело уже с вероятностью, чем больше концентрация субстрата тем больше вероятность того, что фермент свяжется именно с субстратом и образует ES комплекс, а не EI.
1 ответ
Понял, ракмет !


По всей видимости под «невысокими» только низких жирафов подразумевают. Если так, то есть два следующих решения решения:
@tardigrade спасибо за второй метод решения


Решение задачи загрузила по этой ссылке: https://cdn.bc-pf.org/ask/genetics30-06-2021.pdf
скрины тоже приложу на всякий случай





если инверсия произошла, значит участок на 180* перевернется. При кроссинговере, при синапсисе они слипаются в месте центромер, и они обмениваются участками. Теперь все это представьте в голове. У нурхана парацентрическая инверсия. если дальше вы по размышляете, тогда будет вероятность после кроссинговера дупликация хромосом, отсутствие цетромер или же наличие двух центромер и удаление хромосомы.
1 ответ
а кто сказал что третье не верно?))) оно верное. Здесь наблюдается парацентрическая инверсия как говорил ранее. Если произойдет кроссинговер, то он будет неравным. Из за этого может возникновение дупликации одной хромосомы и делецию гомологичной. Надеюсь теперь все понятно))) если ответ понятен и не осталось других вопросов, попрошу поставить “вопрос решен”
Привет!
Для решения этой задачи стоит начать с определенных предположений: задача решаемая только если концентрация маннита в крови и межклеточной жидкости находится в равновесии: после часа нахождения в собаке это вполне возможно. Исходя из этого, мы можем использовать концентрацию меченного альбумина, который из крови выйти не может, чтобы найти объем крови. 50мг введенного альбумина мы делим на 0.031мг на мл в крови и получаем 1612.9мл крови. Используя этот объем, мы умножаем его на 0.29мг маннита на мл крови, чтобы получить часть маннита в крови. Затем мы вычитаем эту массу из введенных 2000мг маннита, и получаем 1532мг, из чего мы также вычитаем 100мг (2000×0.05) потерянные с мочей. Полученная масса – масса маннита в межклеточной жидкости. Исходя из нашего предположения, мы делим ее на концентрацию маннита в межклеточной жидкости = с_маннит в крови = 0.29мг на мл.
Ответом является 4938мл=4.94л, что и есть опция В

На уровне середины указательного пальца, давление равно 30 мм.рт.ст, а на уровне подмышки, давление равно 80 мм.рт.ст. Разница между этими двумя давлениями будет равно pgh. Это довольно логично, потому что в точке подмышки, жидкость(кровь) на высоте h до указательного пальца, будет,грубо говоря, создавать дополнительное давление на кровь в этой точке. Теперь нам остается только перевести мм.рт.ст в Па, и использовать уравнение.
Если 1 Па = 0,0075 мм.рт.ст, то очевидно, что 80-30=50 мм.рт.ст будет равно 6666,667 Па. Теперь, нам остается только воспользоваться формулой, и найти h :
6666,667 = 1060 * 9,8 * h
Отсюда h = 0,64 м = 64 см
Я считаю, что в задаче допущена ошибка, ибо плотность крови = 1060 г/см3 как по мне, абсурд. Тут 1060 либо должно иметь единицу измерения кг/м3 , или же единица измерения должна оставаться в г/см3, но с запятой после единицы.
Возможно, у тебя возникнет вопрос, почему в формуле надо использовать давление в Па. Тут дело в том, что 1 Па = 1 кг/(мс2) , и по формуле думаю понятно, что плотность надо брать в кг/м3 , константу гравитации оставить как есть, т.е м/с2, а высоту брать в м. Если умножить эти единицы измерения друг на друга, как раз так и получится кг/(мс2), или другими словами, Па.
1 ответ
получается там должно быть P = pgh, а не P + pgh? блин, это меня и смутило. но я до конца не верил, что это ошибка, думал, мол респа же, вряд ли такая грубая ошибка будет (типа может быть на другом конце уравнения нужно что-то самому посавить)
ну и про мм.рт.ст я не знал
спасибочки ^
нужно было всё-таки на уроках физики сидеть, там вроде что-то такое проходили. и в итоге я эту задачку на респе не смог решить((((
2 ответа
Вообще, задачу можно было решить и с помощью правой части графика(или как это называется?) не используя мм.рт.ст. Пусть цена деления будет х кПа, и по графику мы видим, что расстояние от n до n+4 кПа равно 3x кПа. Теперь, ты можешь найти цену деления, с помощью уравнения n+4 - n = 3x , x = 1,333 кПа.
Давление в точке подмышки равно 8+2x = 10,667 кПа, а давление в точке середины указательного пальца равно 4 кПа. Разница в давлениях будет равна 6.667 кПа = 6667 Па, что и соответствует моему решению сверху.
про это даже не подумал) мозги у меня, канешна, не любят работать
Если честно, ошибки в задаче кроме плотности я не вижу. Давление должно быть Р + pgh в точке подмышки , это факт. А давление в точке середины указательного пальца равно Р. Стоит отметить, что здесь надо понимать, что мы нашли разницу в давлениях, и эту разницу приравняли к pgh потому что P + pgh - P = ΔР ; pgh = ΔP










