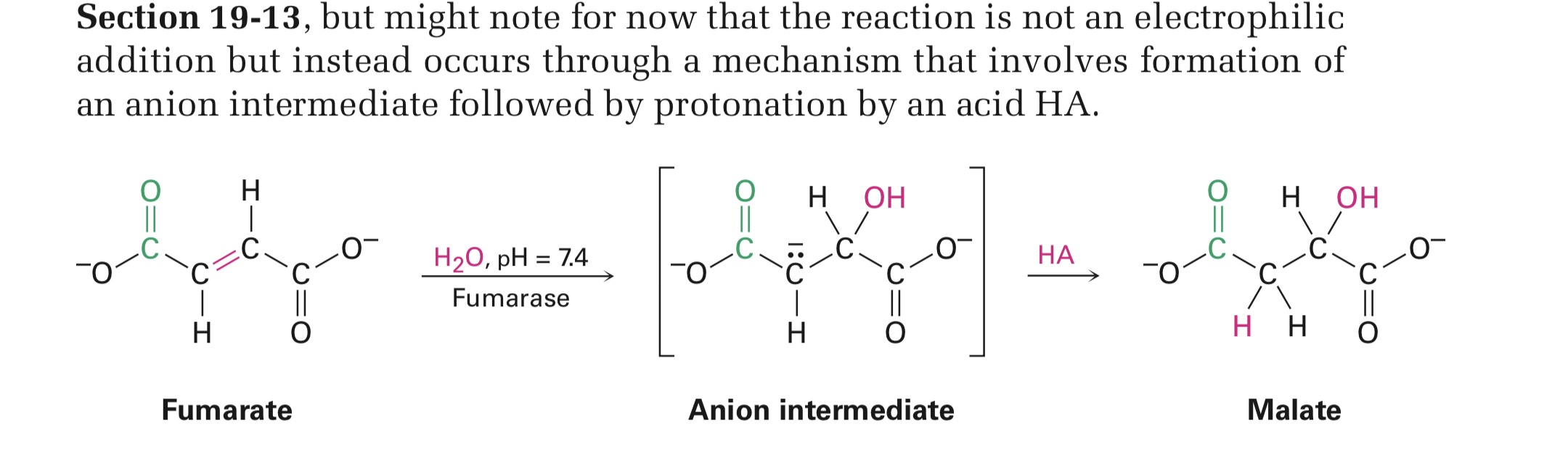
Почему биологические гидратации требуют, чтобы двойная связь была рядом с карбонильной группой?
2)Почему в реакции сверху нету электрофильного добавления как оно обычно бывает, вместо этого сперва добавляется ОН и молекула становится анионом, а затем ее протонируют НА?
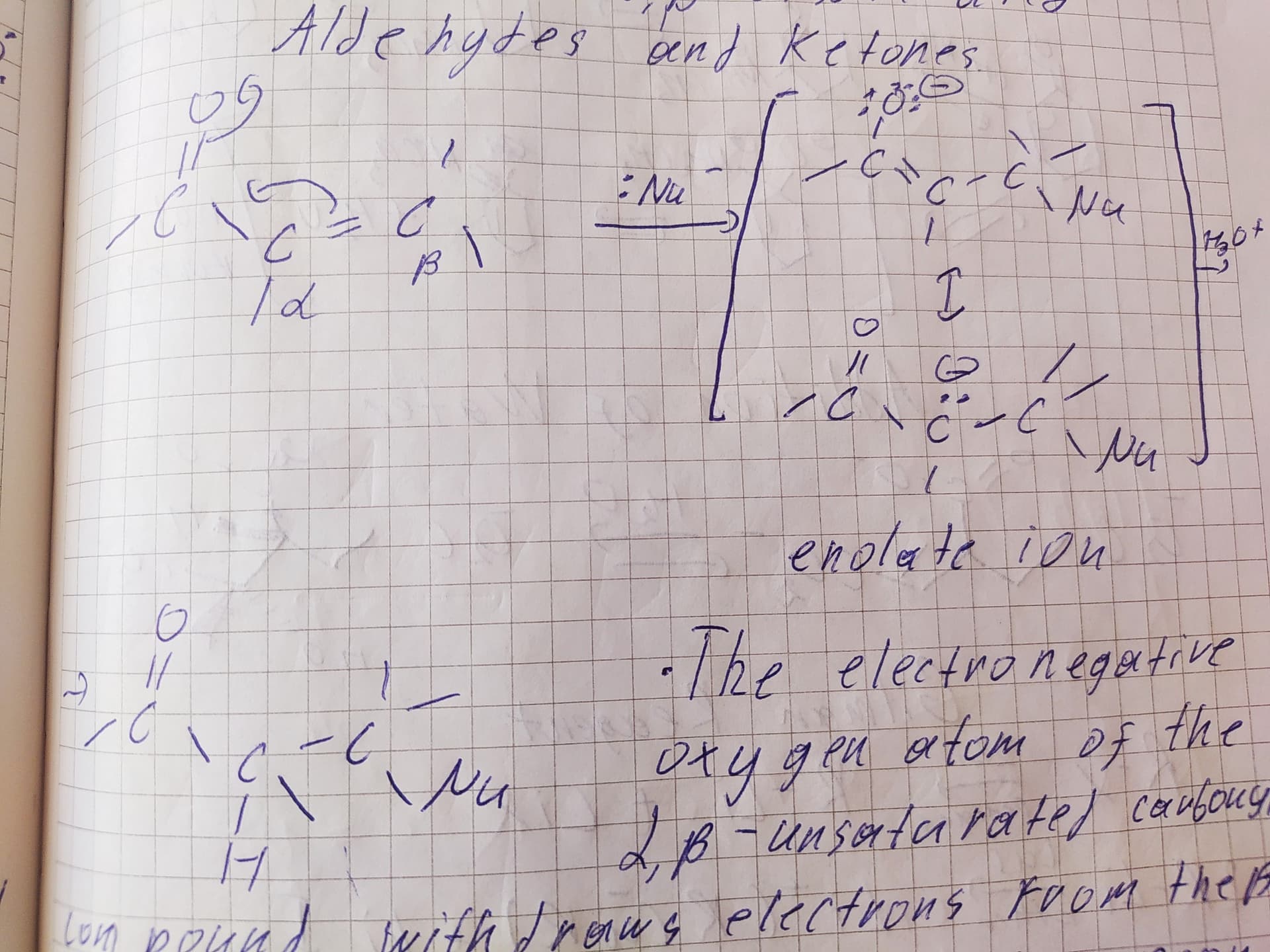
Тут изображено присоединение к сопряженным карбонильным соединениям(то есть где двойная связь рядом с карбонилом). В таких молекулах бета-атом углерода от карбонила является электрофильным(попробуйте нарисовать резонансные структуры этой молекулы), он присоединяет гидроксильную группу(нуклеофил), а на альфа атоме остается отрицательный заряд(образуется енолят-анион), после протонирования получается продукт реакции.
Значит дело в резонансе? Типо мы не можем это контролировать, и так как настоящая молекула это некий гибрид между двумя резонансными структурами, значит молекула в какой-то степени и нейтральна, и имеет отрицательно заряженный кислород с положительно заряженным углеродом, и поэтому при реакциях этот атом углерода скорее является электрофилом. А в этом случае мы игнорируем правило Марковникова?
да, если сложить все структуры то у нас получится что положительный заряд распределен между связанным с кислородом и бета атомами углерода(карты электронной плотности подтверждают). Да, правило Марковникова тут не учитывается
А они требуют? Здесь просто указана одна из реакций цикла Кребса, с участием фумарата.
Потому что эта реакция протекает в активном центре фермента и поэтому музыку выбирают не “общие принципы органической химии”, а качественное и количественное расположение аминокислот в активном центре фермента.
Может так получаться[1], что в активном центре есть нужные аминокислоты, которые быстренько депротонируют воду и происходит присоединение \ce{OH-} с образованием карбаниона.
А может в теории быть и наоборот – сначала идет протонирование, а потом присоединение.
Важно понимать, что белки настолько мощные ребята, что могут создавать внутренние кармашки, в которых среда по своим характеристикам будет разительно отличаться от раствора (иными словами, даже если белок растворен в pH=7.4, где-нибудь может быть кармашек с pH=4 или 9)
Половина МакМурика построена на том, чтобы уйти от догматизма “правил Марковникова” и иже с ними.
Мы не игнорируем правило Марковникова, просто правило Марковникова это короткое обобщение принципов протеканий реакций для определенных субстратов. Как только у нас другие субстраты – сразу же это “правило” перестает быть актуальным.
В данном случае, если вы захотите нарисовать электрофильное присоединение так, чтобы гидроксогруппа оказалась на альфа атоме, вы должны будете нарисовать предшествующий карбокатион, где положительный заряд находится на атоме углерода в альфа положении. Надо ли говорить, что такой карбокатион будет менее стабильным, чем карбокатион, где положительный заряд находится на бета углероде?
Стабильность карбокатиона самое главное. Иногда стабильность карбокатиона совпадает с правилом Марковникова, иногда нет – но это проблемы правила Марковникова, а не карбокатиона.
Если же вы хотите нарисовать нуклеофильное присоединение, то тут механизм, который показал @elprimo. Опять же, нарисуйте с другой региохимией и получите карбанион с отрицательным зарядом на бета углероде. Надо ли говорить, что такой карбанион менее стабилен, чем тот, где отрицательный заряд на альфа положении?